Neogene relicts(新第三紀遺存種、新成紀遺存種) 前 千葉大学 園芸学部 岡上伸雄 I. Stenophora 節 Burkill (1960)1)は、化石や大陸移動などと考えあわせ、rhizomatous type がより原始的であり、おそらく Stenophorans は Eocene (始新世; 第三紀の前半の2番めの epoch) には北半球に広く分布し出していたが、乾燥化に続いて寒冷化も進んだ Miocene (中新世; 第三紀の後半の最初の epoch) を経て今につながる第四紀になる頃にはおもな分布域は湿潤な東アジアのみになり、一方 tuberous type は水分や粘質物を多く含むために、Miocene に繁栄しだした、と推定した。 1) Burkill, I. H. (1960): The organography and the evolution of Dioscoreaceae, the family of the yams. Journal of the Linnean Society (Botany), 56, 319-412. 2) 分布域が北の種から、①ウチワドコロ、②オニドコロ、③タチドコロ、④キクバドコロ、⑤ヒメドコロ、⑥カエデドコロ、⑦イズドコロ、⑧シマウチワドコロ、⑨ツクシタチドコロ; 以上9種。 もし、コシジドコロを加えると(④と⑤のあいだに)、10種類となる。 |
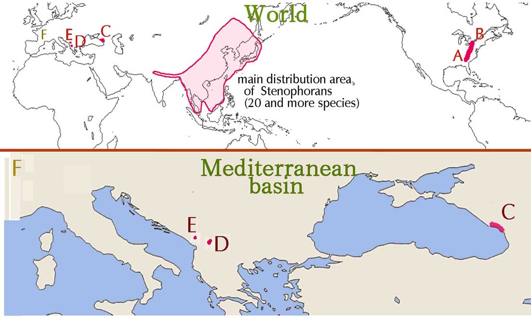
II. Tertiary relicts → Neogene relicts (第三紀遺存種 → 新第三紀遺存種 または 新成紀遺存種) このStenophora節の特徴的な形態(地下肥大器官が根茎、葉は心形単葉、茎は左上がり巻き、雌花は花後反転して子房が上を向く、など)をもつ4種と1formが、東アジアから遠く離れた北米東部、小アジア、バルカン半島などの狭い範囲に隔離分布をしている。これらの分布地を、Burkill (1960)1)の地図を少し改変して下に示す。上半分の世界地図には東アジアの20余種の全体の分布域とともに、隔離分布している種(A-E)の分布地域のおおよそを示し、下半分には地中海域の詳細を示した。 FはStenophoraの葉の化石3)がEoceneの地層から出土したフランスのパリ盆地である。この化石種は Dioscoroides Lyelli (Wat.) Fritel と命名されている。 3) Potonie, H. (1921): Lehrbuch der Paläobotanik, Zweite Auflage von W, Gothan. Gebrüder Borntraeger (Berlin). のp356, Fig.279.
ところで、しばらく前に地質時代の名称が変更され、Tertiary (第三紀) というくくりの period は廃止されてこの語は用いられなくなった。Tertiary という period は前半と後半の二つに、即ち Paleogene (古第三紀、旧成紀) と Neogene(新第三紀、新成紀)とに分けられた4)。 Burkill1)によれば、Stenophora のうち遺存的なメンバーが生き残ることになったのは、Miocene の時代からとされているため、この epoch を含む period である Neogene の名をつけて、今後はこれらの遺存種を Neogene relicts (新第三紀遺存種 または 新成紀遺存種) と称するのが適当と思われる。 なお、Paleogene と Neogene のより適切な日本語訳については石渡の提案がある5)。 4) ICS: International Chronostratigraphic Chart, v2015/01. International Commission of Stratigraphy. http://www.stratigraphy. org/index.php/ics-chart-timescale
5) 石渡 明 (2006): 新生代の紀の名称として「古獣紀・新獣紀」の提案。 http://earth.s.kanazawa-u.ac.jp/ishiwata/kojuki.htm#top
6) Terui, K. & Okagami, N. (1993) Temperature effects on seed germination of East Asian and Tertiary relict species of Dioscorea (Dioscoreaceae). American Journal of Botany 80, 493-499. (1993)
7) García, M. B., Antor R. J. & Espadaler, X. (1995) Ant pollination of the palaeoendemic dioecious Borderea pyrenaica (Dioscoreaceae). Plant Systematics and Evolution 198, 17-27.
|
III. 北米東部 D. quaternata は、五大湖の少し南を分布の北限としている。オハイオ州内の詳細な分布図9)によると、この北限はウィスコンシン氷河の氷床の南端から南方へ最大で幅が約200kmあったと思われているツンドラ状の地域8)の南縁の線に一致しているように見える。分布の南限はアパラチア山脈の南端、フロリダ州にまでかかる。一方、 D. villosa の分布の北限はツンドラの南縁を北へ抜けて五大湖の北岸 (カナダ) にまで達している。オハイオよりも南の数州では両種は共存している。 8) 棚井敏雅 (1992),"アメリカ大陸の森林の歴史" In 赤澤ら編 アメリカ大陸の自然誌 1 アメリカ大陸の誕生.岩波書店、 p117-180.
9) Braun, E. L. (1967): "The Monocotyledoneae, Cat-tails to Orchids" In Braun, E. L. (ed) The Vascular Flora of Ohio Vol. 1, Ohio State Univ. Press, p392. |
| D. quaternata Gmelin (地図のA) : Pennsylvania 州で採取し、日本で栽培している株を下に示す。 | ||
 |
 |
 |
| 〔写真左〕 D. quaternata (仙台市青葉区で栽培、5月はじめ)。 葉脈の数が、Stenophora の東アジアの種や他の遺存種 (9 -11本) に比べてやや多く、13-15本前後もある。 〔写真中の下段〕はオス株の最下位の節で、5葉が輪生している。下位の数節が輪生葉 (3−7枚程度) をつけることは、程度の違いはあれ (葉の枚数や、葉の高さのわずかな差などに) 、Stenophora の Neogene relicts に共通する性質である。 〔写真中の上段〕に、下から2番めの節を示した。6葉が輪生しており葉腋からオス花序が伸びだしている。シュートは先端生長を続けるため、この節は地上30-50cmほどの位置にとどまり、その高さでオス花は咲く。なお、オス花序はこれより上の数節にもつく。オス花序は、地下器官から伸び出したばかりの shoot に既にみてとれる。 下位の節からオス花がつくことは、そしてまた下位の節に葉が輪生することは、輪生する葉の枚数は少ないものの、日本の九州南部に固有な Stenophora であるツクシタチドコロにも時おり見られることがある。 一方メス株では、花序はだいぶ上の節にしか着かないため、メス株とオス株とが並んでいる時でも、下位の節の高さではメス花とオス花とが接近して咲くことはない。 〔写真右の上段〕はメス花 (仙台市青葉区で栽培、5月上旬) 。6枚の花被は外側 (12時、4時、8時の位置) と内側の3枚ずつに わずかの違いがあり、区別できる。3本の雌蕊はまとまっており柱頭は3本とも二股に分かれている。 〔写真右の下段〕はオス花 (千葉県八街市で栽培、5月上旬) 。花被の区別はメス花に同じ。6本の雄蕊のうち、外側の花被 (12 時、4時、8時) の付け根から出ている3本が、わずかに発達が早い。退化雌蕊 (pistillode) は花の底から明瞭に盛り上がっている。 花の中心から花被の付け根のあたりまで蜜が覆っている。 |
||
| D. villosa L. (地図のB) : 上の種よりも少しだけ北に分布している。上の種に比べて花がやや小ぶりでカプセルと種子も少し小さいこと、下位の葉の輪生の程度が低い(葉の枚数が少ない)こと、オス花序はより上の節から出ること、葉脈の数が多くないこと、花期が少し遅いこと、オス花に弱い芳香があること、などの点以外は、上の種に酷似している。 | |
 |
 |
〔写真左〕 D. villosa のメス花序 (メス花は少しふくらんだ子房を下位にもっている) と、花を密につけたオス花序が見える。メス花序もオス花序もおおよそ10節め以上につくため、両株が並んでいるときには、10節め以上では両花序が混在する状態になる。メス花序はツルの先端近くの節にまで着く。千葉県松戸市で栽培。6月初旬。
10) Prain, D. & Burkill, I.H. (1936). An account of the genus Dioscorea in the East. Part I. The species which twine to the left. Annals of the Royal Botanic Garden, Calcutta 14(1): 1-210. plate 1-84. |
|
| IV. 小アジア
コーカサス山脈とトルコに近いメスヘティ山地とのあいだ、いわゆるコルヒダ低地 (Kolkhida lowland) は、黒海の最奥部に接しており、低気圧が到達しやすく、夏期にもある程度の降水がある。コーカサスを北側にして温暖な地域であり、Fagus orientalis Lipsky (オリエントブナ 或いは コーカサスブナ) が分布しており、また、茶やミカンの栽培も行われている。 D. caucasica Lipsky (地図のC) : コルヒダ低地の北半分の、少しだけロシアに入ったソチの南から、ジョージア (Georgea、グルジア) のアパジア (Apkhazia、アプハジア) 自治共和国の首都のスフミ (Skhumi) 付近まで、黒海沿いの長さ約100Km、幅20Kmほどの範囲にこの種は分布している10)。Fagus orientalis とともにコルヒダフロラの代表的なメンバーであり、両者ともウクライナの V. I. Lipsky により、19世紀末に発見記載された。 11) Gitsba, D. K. (1972). Distribution of Dioscorea caucasica Lipsky and methods of rational utilization of this species. Rastiel Resursy 8: 577-587. |
|
 |
 |
| 〔写真左〕 オス株。下位の葉が3 - 4枚輪生している。花序はオス株もメス株も上位の節に着く (仙台市青葉区で栽培 7月中旬)。 〔写真右〕 メス株。カプセルは花後急速に大きくなり完成時の大きさに達するが、中の種子はまだ小さなことが透けて見える (7月中)。この後約2ヶ月ほどかかって、種子は完成時の大きさになる。 |
|
| V. バルカン半島
バルカン半島の西部のディナールアルプスは北西から南東に長い。隣接する地域とともに、氷期にも生き延びれるような場所が多かったためか、東アジアの植物に関連する遺存種が生育しているスポットが多い。おおまかに見ると地中海性気候で夏季の降水は少ない地域だが、局地的に特異な地点が諸所にある。旧ユーゴスラビアの観光案内パンフレットには、民族が入り組んでいるのと同じように気候も入り組んでいて多様な環境があるために自然が豊かである、とあった。 D. balcanica Košanin は、第一次世界大戦の直前のバルカン戦役に従軍してアルバニア前線に駐屯していたセルビアの植物学者 N. Košanin が、国境の山岳地帯のアルバニア側の中腹から裾野にかけての一帯で1913年に、先ず発見し記載した12)。その後1960年に、この一帯の北に隣接する地域で新たな生育地が見出され分布域はコソヴォ (当時はセルビア) 内で少し広がった (地図のD)13)。 一方、Košanin 門下の Marie Zafirović が、上記の地の中心から北西に約140Kmほど離れたモンテネグロで同種と思われる植物を、1920年代末期に発見している (地図のE)14)。 12) Košanin, N. (1914) Dioscorea balcanica Kos. n. sp. Österreichische botanische Zeitschrift 64: 37-39.
13) Lakusic, R. (1967) Einige Resultate oecologischer Untersuchungen an der Art Dioscorea balcanica Kosanin. Angewandt Pflanzensociologie 18/19: 47-53.
14) Košanin, N. (1929) Nachträge zur Dioscorea balcanica Koš. Bull. Inst. Jard. Bot. Univ. Belgrad, 1: 199-203.
15) Simic, M & Antonovic, A. (2007): A new view on the structural pattern of the Metohiya Basin and its margin: a preliminary note. Annales Geologiques de la Peninsule Balkanique, 68, 29-38.
16) Zogovic, D. (1993) Storage watertightness problems and technical solutions in the highly developed karst area of the Niksic polje, Yugoslavia. Proceedings of the Antalya Symposium and Field Seminar, October 1990. IAHS publ. no. 207: 85-93.
D. balcanica Kosanin North Albanian-Kosovan form (地図のD): タイプ標本17)が採られた集団が生育している地域は、アルバニア北部の都市クカス (Kukes) の近く、山岳地帯の麓の村ビチャイ (Bicai) から、ジャリツァ山 (Gjallica) とコリトニク山 (Koritnik) の中腹を経て、メトヒャ盆地からアルバニアに流れ入るドリン川 (Beri Drim) を挟んで向かい合うパストリク山 (Pastrik) の中腹を辿って、国境を越えたコソヴォ共和国 (またはセルビアのコソヴォ・メトヒャ自治州) 南西部の都市、旧ユーゴスラビアの中でももっともオリエント風の雰囲気を残すプリズレン (Prizren) の西郊に至るまで、約20数km ×10数km ほどの範囲である13)。 17) Vukojičić, S. et al. (2011). University of Belgrade Herbarium--treasury of data and challenges for future research. On the occasion of the 150th anniversary of University of Belgrade Herbarium (1860-2010). Botanica Serbica 35, 163-178 のFig.8.
18) Sustainable Land Use Management and Conservation of Biodiversity in Dragash/š Municipality. UNDP in Kosovo.
|
 |
| 〔写真上〕 アルバニアの北東部クカス県の山岳地帯 (2002年9月8日)。山の向こうはコソヴォ、右奥方向はマケドニア。山の中腹から上には雲がかかることが多く、驟雨性の雨も多い。D. balcanica は、おもに雲霧によって涵養されていると思われる低木にからまって生育している。ここに見える範囲では、燃料として伐採され難いこともおそらくかかわって、標高が高いほど樹高も高い。森林監視官によると、現在は低木林になっている標高でも、数十年前までは大木が生えていた跡があるのだとのこと。山麓のエントツ (画面右端) はかつての銅の精錬工場のもの。ここからの排煙によっても植生が大きな影響を受けたようだとのこと。 |
 |
 |
| 〔写真左〕 North Albanian-Kosovan form の生育範囲 (地図のD) のコリトニク山中で木にからまって生えている D. balcanica。 潅木林の林縁ではなく、林内の木に絡まって生育している株が多い。 茎葉は十分に成長している個体が多く、メス花も着いているが、果実にまで至っているものは少ない。 しかし、ここで採取した株を平地で栽培するとメス花のほぼ全数が健全な種子を形成するまでに至る。 この地点で見られる株の外部形態は、タイプ標本17)のものとよく一致している。 ところで、Lakusic (1967)13) には、Dの生育地域の北端に接する都市プリズレンの西の郊外 (標高が低い箇所、本文参照) に生育している株のものとして、茎の断面と葉の形の図が描かれている。しかし、その形態は、Kosanin のタイプ標本17) のものとはだいぶ異なっている。 Lakusic が描いたプリズレンの株はDの地域内の変異株の可能性はある。 一方、Lakusic のプリズレンの株の茎葉の図は D. caucasica の茎葉に酷似している。何らかの経緯により D. caucasica と取り違えられているという可能性もあろう。 Dの生育地域の大部分は政治的問題をはらんだ国境の危険な地帯であった期間が長く、地雷もたくさん埋められた地域であったため詳細な調査が進んでおらず、Dの生育地域内の小集団の調査が必要であろう。 〔写真右上〕 隣の個体までの距離が遠く、疎に分布している。この写真の中には、矢印の先の画面中央に一株だけ。 |
|
〔写真右〕 メス花。メス花の花被はオス花に比べるとやや短い。6本の退化雄蕊のうち、外側の花被の付け根から出ている3本のほうがわずかに発達が早い。
|
 |
〔写真右〕 オス花。雄蕊の3本ずつに発達速度の違いがあることはメス花に同じ。横から見ると、花の中央部のカップ状の部分の深さが、北米の種に比べるとやや深い。千葉県八街市で栽培中の株のもの (7月22日)。 |
|
| D. balcanica Kosanin Montenegrin form (地図のE): この集団は、モンテネグロ第二の都市のニクシチ (Niksic) の市内の Gymnasium の近くの大きな泉の近くで M. Zafirović が発見し14)、直径20km足らずの範囲 (約350km2) にだけ分布していることが知られた13)。Stenophoraの現生種としては、もっとも西に分布している集団である。この分布域の大部分は、大きなニクシチポリエから成っており、1988から2002年にかけての調査では、このポリエの縁に該当する箇所の、なかでも撹乱を受けたと思われる箇所 (道路開設、工場建設、宅地開発、ダム工事などによる) に多く見られた。 それらの生育箇所では、種子から1 - 2年前に発芽したと思われるような個体もたくさん見られた。 おもな生育箇所の標高は620 - 680m程度である。 この直径20km足らずの範囲内では、Lakusic が述べているように13)、各生育箇所の群落間では隔離は生じていないと思われる。 | |
 |
|
| 〔写真上〕 Niksic市の西方の郊外。牛の放牧地や小規模な畑地に隣接する石灰岩のくぼ地の斜面や底の土壌がたまっているところに、潅木にからまってこの植物は生えている。 |
|
 |
|
| 〔写真上〕 画面の左端から1/3 ほどのところに写っている木の上のほうに、赤みがかった色の数個のカプセルを付けたツルが絡まっているのが見える。この株の根元は画面右端下部。葉の形は、1914年に北アルバニア-コソヴォ地域で採取されたタイプ標本17) の葉によく似ている。左端の林内には石灰岩塊と、咲き出したピンクのシクラメンの花が見える (2002年9月13日)。前掲のアルバニアのコリトニク山の集団とは違い、種子をもつたくさんのカプセルをつけた株が多い。5月末頃に形成されはじめたカプセルが成熟するまで4ヶ月はかかる。からまれている木はおそらく Carpinus orientalis。 コリトニク山の集団 (North Albanian-Kosovan form = 母種) とニクシチの集団 (Montenegrin form) の個体は、よく似た形態をしていて容易には区別できない。しかし、これら二者を並べて栽培して複数年にわたって観察すると、さほど大きくはないが一定の生長差があることは認められる。 D. caucasica の項で触れたのと同様に、この地域でも、地中海域のヤマノイモ科の種である Tamus communis と D. balcanica とが同じ場所に絡まりあって生育しているのをよく見かける。Tamus communis と D. balcanica の種子は、自然状態では両者とも春のほぼ同じ時期に発芽することが多いが、両者の種子の休眠の制御機構はまったく異なっている。また、種子の胚乳中のトリアシルグリセロール分子種組成も、両者の間で対照的に異なっている。 |
|
| 〔写真右〕メス花。 |
 |
〔写真右〕オス花。
|
|
上に述べてきたように、北米東部、小アジア、バルカン半島の Neogene relicts の間の形態上の違いは、東アジアのStenophora 節の種の間の違いに比べるとわずかであり、Neogene relicts の生長の仕方もお互いに似ていて、区別は簡単ではない。さらに、種子の休眠性、発芽や芽生えの成長時に消費される貯蔵脂質のトリアシルグリセロールの分子種組成のような性質も、東アジアの種が有している性質とは対照的に異なっていて、遺存種のあいだでは違いは少なくほぼ一致している。遺存種が一致して持っているこのような性質の多くが、隔離前の性質である可能性を考えている4)。 Stenophorans は、東アジアにおけるおそらく温度に関連する要因による南北分布と、ユーラシアと北米をつなぐ広い地域の両端にわたる歴史的経緯がかかわる遺存種の東西分布とを持つ多年生草本の種の系列であり、さまざまな観点による研究がなされているが、ここでは以下に分子遺伝学的な知見を簡単に紹介する。 |
|
VI. 分子遺伝学の知見 19) Kawabe, A., Miyashita, N.. T. & Terauchi, R. (1997). Phylogenetic relationship among section Stenophora in the genus Dioscorea based on the analysis of nucleotide sequence variation in the phosphor glucose isomerase (Pgi) locus. Genes Genet. Syst. 72: 253-262.
20) 河邊 昭 (1997). Pgi 遺伝子座のDNA変異に基づくヤマノイモ属ステノフォラ節の系統関係. Dioscorea Research 1, 59-62.
21) 丹野憲昭、M.B.Garcia ら (1997). ヤマノイモ科 (Dioscoreaceae) の内生ジベレリン. Dioscorea Research 1, 75-83
〔上記の遺存種の採取や現地観察は、在東京の各国大使館の許可を得るか、各国の然るべき機関を介して行った。 花の外部形態の写真は随時改訂を行う予定。〕 |